Il modello aggiornato dei metabolismi energetici

(Aggiornato al 22/05/2023)
Questo articolo tratta dei marginal gain (cioè quei piccoli dettagli che possono fare la differenza) che si possono ottenere grazie ad una conoscenza più accurata dei metabolismi energetici.
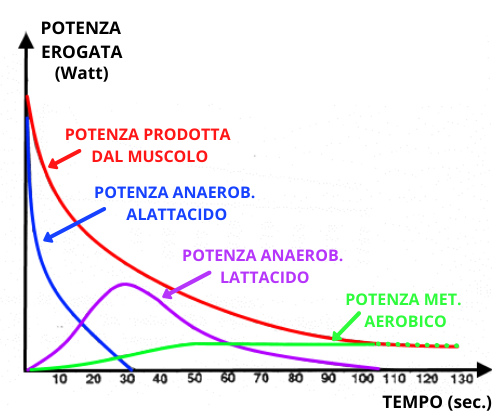
In rete ed in bibliografia internazionale è possibile trovare una moltitudine di articoli che riguardano il funzionamento dei metabolismi; la quasi totalità di questi si basano su presupposti sperimentali su dati ottenuti da biopsie e Risonanze magnetiche nucleari (RMN) con campionamenti abbastanza ampi come 5” e più. Questi metodi d’indagine permettono di fare una fotografia istantanea di quelli che sono metaboliti in quel momento, che confrontati con la situazione a riposo (o intermedia) permette di ipotizzare il funzionamento della biochimica muscolare. Fino a circa 20 anni fa questi erano i metodi d’indagine principali che ci permettevano di estrapolare il funzionamento dei metabolismi energetici; nell’immagine sotto potete vedere uno schema semplificativo.
Ma questi concetti sono validi tutt’oggi? I moderni metodi d’indagine (come la Risonanza Magnetica Nucleare con campionamento al millisecondo) hanno cambiato la nostra conoscenza dei metabolismi? Quali applicazioni pratiche ne possiamo dedurre?
Questo post è dedicato a tutti quegli addetti ai lavori che sono interessati ad approfondire questo argomento, come laureati in Scienze Motorie, diplomati ISEF e tutti coloro che conoscono la biochimica dell’esercizio.
Partiamo da un presupposto: la Risonanza Magnetica Nucleare con campionamento al millisecondo rispetto ai vecchi metodi d’indagine, può offrire una mole di informazioni più vasta ed approfondita di tutto quello che accade nel muscolo durante l’esercizio fisico. È ovvio che non rivoluziona completamente la vecchia visuale dei metabolismi (che per certi versi rimane corretta), ma le informazioni che si possono avere permettono di ottenere dati che prima non si avevano, con ripercussioni importanti anche in metodologia d’allenamento.
In questo articolo andremo ad analizzare la Teoria dello Shunt del glicogeno per poi vederne le diverse ricadute applicative; approfondiremo come il lattato non sia la “causa” della fatica, ma il “fulcro del metabolismo”. Vedremo poi come il microbiota intestinale abbia un ruolo estremamente funzionale anche nei confronti della performance (vedi il “Caso Veillonella”). Non solo, analizzeremo le ultime ricerche in ambito di modificazione dietetica per il miglioramento della performance di endurance e nuove tipologie di integratori a base di carboidrati. Per ultimo (ma non meno importante) citeremo alcune ricadute applicative della Teoria dello shunt del glicogeno in ambito metodologico.
Come potete notare, l’articolo è parecchio lungo, quindi consiglio di leggerlo in più riprese; vi lascio al sommario e di seguito al primo capitolo. Buona lettura!
INDICE DEI CONTENUTI
“Shunt del glicogeno”: ipotesi o rivoluzione?
Secondo il vecchio paradigma dei metabolismi, maggiore è l’intensità dell’esercizio e maggiore è l’attivazione dei metabolismi anaerobici; questo perché il metabolismo aerobico non è in grado di supportare l’intera produzione di ATP quando al muscolo è richiesta una potenza elevata. Non solo, anche nei primi secondi dell’esercizio (soprattutto se intenso) il contributo dei metabolismi anaerobici sarebbe maggiore, proprio perché quello aerobico ha una certa “inerzia” nel mettersi in moto.
Quello che è stato scoperto con la RMN con campionamento al millisecondo, ha permesso di approfondire ulteriormente quello che accade nel muscolo nei vari istanti della contrazione muscolare; infatti, non si tratta più di “secondi”, ma di “millisecondi”.
Con le sue 2 revisioni (quella del 2000 e del 2005) Shulman ha riassunto quelli che sono stati i dati ottenuti dalle RMN (campionamento al ms) su animali ed uomini, postulando la teoria dello Shunt del glicogeno* che esporremo di seguito.
* Importante: la teoria dello Shunt del Glicogeno è desunta da indagini effettuate su sforzi di intensità limitata e di lunga durata.
Durante i primi 15 ms (millisecondi) di una contrazione muscolare non massimale (come camminare) l’ATP è ripristinata prevalentemente dalla Fosfocreatina, che dopo 8 ms viene già dimezzata. Dal 15° ms (circa) fino al 100° ms della contrazione (ipotizzando che questa possa esserne la durata) l’ATP viene ripristinata principalmente dalla glicogenolisi/glicolisi, che non solo permette di sostenere la contrazione muscolare, ma consente (durante la contrazione stessa) di ripristinare parte dalla Fosfocreatina (vedi immagine sotto)
Ricordo che per semplificare, per glicogenolisi/glicolisi si intende l’insieme delle 2 vie metaboliche che permettono di ottenere inizialmente glucosio6-P dal glicogeno (glicogenolisi) e poi di ripristinare 3 ATP e produce 2 molecole di lattato (glicolisi).

Quello che emerge dalla teoria dello shunt del glicogeno (che si basa su dati sperimentali) è che l’energia necessaria ad ogni contrazione muscolare (anche a basse intensità) è fornita dalla fosfocreatina ed in gran parte dalla glicogenolisi/glicolisi. Questo, perché il fabbisogno energetico per la contrazione è talmente elevato da non poter essere sostenuta in buona parte dal metabolismo aerobico (anche a basse intensità).
Quindi, rispetto a quello che è il vecchio paradigma dei metabolismi energetici, non si parla più di “secondi di sforzo”, ma di “millisecondi” all’interno di ogni singola contrazione muscolare.
Ma a cosa serve alla cellula il metabolismo aerobico?
Serve principalmente per fornire l’energia per ripristinare il glicogeno durante la fase di rilassamento muscolare (vedi immagine sotto). Mi spiego meglio: ad ogni contrazione muscolare, viene consumata all’incirca 1 mM/g (millimole su grammo) di glicogeno; partendo dal presupposto che la concentrazione di glicogeno nel muscolo è in media 70 mM/g, se non fosse ripristinato durante la fase di rilassamento, dopo 70 contrazioni sarebbe completamente esaurito.
All’interno dell’energetica della contrazione/rilassamento muscolare, il metabolismo aerobico ha la funzione principale di fornire l’energia necessaria (2 ATP) per aggiungere il glucosio proveniente dal sangue al glicogeno.

Questo avviene perché il tempo di rilassamento della singola fibra muscolare in sforzi di bassa entità è molto più lungo, in quanto queste non entrano in funzione tutte durante la contrazione; esiste un turn-over (questo fenomeno si conosce da tempo) che recluta a “rotazione” le fibre in base all’entità dello sforzo, al grado di affaticamento, ecc.
È quindi ragionevole ipotizzare come la maggior parte del glucosio che entra nella fibra muscolare non venga ossidato a scopo energetico, ma vada a ripristinare le unità glicosidiche del glicogeno. Allora quali sono i substrati principali del metabolismo aerobico nella fibra?
Semplice, il lattato principalmente (Brooks 2020), poi in misura minore gli acidi grassi e gli scheletri carboniosi degli aminoacidi utilizzati a scopo energetico.
Ma facciamo ora un breve riepilogo per avere le idee più chiare (potete vedere una sintesi semplificata nell’immagine sotto): durante la contrazione muscolare la glicogenolisi/glicolisi provvede a ripristinare una gran parte dell’energia necessaria (riformare ATP). Il sistema della fosfocreatina aiuta nei primi millisecondi dello sforzo, mentre il metabolismo aerobico probabilmente fornisce un contributo minore alla contrazione. Infatti, quello che è importante capire, è che le indagini con la RMN hanno evidenziato come durante la contrazione sia necessario ripristinare l’ATP ad una velocità tale che non può essere sostenuta dal metabolismo aerobico, ma deve essere coperta principalmente della glicogenolisi/glicolisi (anche in sforzi di bassa intensità). Per questo motivo, una buona parte del glucosio che entra nella cellula (durante l’attività muscolare) va a formare le unità glicosidiche del glicogeno.
Questo è stato dimostrato (sempre in sforzi di bassa intensità e lunga durata) all’interno di fasi dell’esercizio in cui la concentrazione di glicogeno rimaneva pressochè costante; grazie all’utilizzo della RMN al Carbonio 13 (13C RMN) è stato visto (dopo qualche ora dall’inizio dello sforzo) come tra il 25-100% del glucosio facente parte del glicogeno non era quello presente prima dell’inizio dell’attività.
Mi spiego meglio con un esempio: ipotizziamo che il glicogeno della fibra sia una grande costruzione di Lego, e che le unità di glucosio che lo compongono siano i mattoncini. Dopo qualche ora di attività, questo Lego sarà leggermente inferiore rispetto all’inizio, ma una parte considerevole (dal 25 al 100%) dei mattoncini che lo costituiscono, non saranno gli stessi dell’inizio.
Ovviamente non si hanno attualmente un numero di dati sufficienti per comprendere con precisione in che misura la glicogenolisi/glicolisi influenzi la produzione di ATP necessaria per la contrazione di bassa intensità: può essere il 90%, come il 75%, oppure il 60%, ma comunque è una parte consistente. Il restante è fornito dal metabolismo aerobico e dal sistema della fosfocreatina.

Durante la fase di rilassamento invece, l’energia fornita dal sistema aerobico contribuisce a ripristinare in via definitiva la fosfocreatina, contribuisce a fornire energia per riformare glicogeno grazie al glucosio proveniente dal sangue (glicogenosintesi) e a produrre l’ATP necessario per il mantenimento dell’omeostasi e delle altre funzioni cellulari (funzionamento pompe ioniche, ripolarizzazione, ecc.).
Dove va a finire tutto quel lattato?
Una volta compreso come esista un continuo turn-over di glicogeno durante le fasi di contrazione/rilassamento, facciamo un attimo il punto sulla funzione del lattato; come abbiamo visto sopra, per ogni molecola di glucosio proveniente dal glicogeno, otteniamo 2 molecole di Piruvato, che vengono trasformate in Lattato; infatti, se a riposo il rapporto tra le concentrazioni di lattato e piruvato è 10, sottosforzo supera i 500. Questo rafforza ancora di più la teoria dello Shunt del Glicogeno, dimostrando come una grandissima parte del glicogeno venga trasformato in lattato.
Mentre una volta si credeva che il lattato fosse la causa della fatica, oggi sappiamo che non è così; lo possiamo considerare come un “testimone” (cioè è presente in elevate concentrazioni quando il muscolo è in alcuni stati di affaticamento), ma non la causa; nei nostri 2 post dedicati all’argomento (primo e secondo) potete trovare il modello tridimensionale della fatica.
Una molecola di lattato, una volta riportata a Piruvato ed ossidata, permette di “ottenere” 18 molecole di ATP grazie alla metabolismo ossidativo (mitocondrio). Secondo la teoria dello Shunt del Glicogeno, il 10% del lattato prodotto dalla glicogenolisi/glicolisi durante la contrazione, può essere utilizzato per fornire l’energia necessaria a risintetizzare il glicogeno durante la fase di rilassamento.
Ma dove va a finire il restante 90%?
Durante sforzi di lunga durata e bassa intensità, dopo i primi minuti di assestamento, la concentrazione di lattato nel sangue rimane costante, segno che esiste equilibrio tra la parte prodotta e quella smaltita; nell’immagine sotto potete vedere quello che può essere il destino di questa molecola all’interno dell’organismo (Brooks et al 2018).
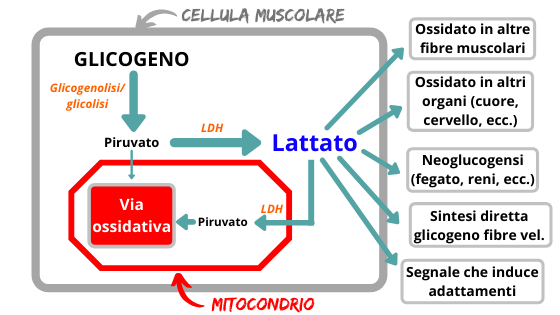
Oggi sappiamo che questa molecola può avere sia funzioni energetiche che glucogenetiche, oltre ad essere un “segnale” in grado di influenzare la secrezione di ormoni (come il GH) e tante altre funzioni cellulari e tissutali. Per questo motivo, dal punto di vista della metodologia dell’allenamento riveste sempre più interesse il “come” viene metabolizzato, affinchè possa essere utilizzato al meglio dal nostro organismo al fine di ottimizzare la performance. A mio parere, è molto importante comprendere la funzione glucogenetica e il rapporto con il microbioma intestinale; nei prossimi capitoli potete trovare gli approfondimenti.
Altra molecola il cui comportamento è stato in parte rivisto è il glucosio; una volta si credeva che durante lo sforzo questo entrasse nella cellula per essere immediatamente ossidato a scopo energetico. Grazie alla teoria dello shunt del glicogeno invece, ora sappiamo che una parte consistente di esso va a formare le unità glicosidiche del glicogeno, poi successivamente utilizzate a scopo energetico. È stato dimostrato (Shulman 2005) che quando la concentrazione di glicogeno nella singola fibra muscolare scende sotto un certo livello, questa va incontro ad affaticamento (cioè non riesce ad erogare la stessa potenza di prima); ne deriva che è fondamentale l’ingresso di glucosio nella cellula per mantenere il turnover di glicogeno (sintesi/degradazione) costante. Ma se nei muscoli è stoccato in media 200-400 g di glucosio (sottoforma di glicogeno), come si riesce a mantenere il turnover se nel sangue ce ne sono solamente 5g?
Una buona parte viene immessa nel sangue dal fegato, grazie alla neoglucogenesi ed in parte dal glicogeno epatico (75-100g), come una parte può essere ingerita sottoforma di integrazione a base di carboidrati. La neoglucogenesi, a questo punto, riveste un ruolo estremamente importante nell’esercizio fisico, perché ha lo scopo di sintetizzare glucosio da altri substrati, tra i quali il lattato.

Non solo, altro aspetto fondamentale (negli sport di endurance) riveste la capacità di rendere disponibile velocemente i carboidrati ingeriti, alla cellula muscolare; è allenabile questo processo? Che caratteristiche deve avere un integratore a base di carboidrati per ottimizzare questa funzione? In che modo, con l’allenamento, si può influenzare la neoglucogenesi? Risponderemo a tutte queste domande nei prossimi paragrafi.
Cosa non dice la teoria dello shunt del glicogeno sul metabolismo aerobico
La comprensione del funzionamento e dell’integrazione dei vari metabolismi energetici coinvolti nella contrazione muscolare è ancora ben lontana dall’essere definitiva; quello di cui si può essere certi, è che attualmente, grazie all’uso della RMN con campionamento al ms, ne sappiamo più di quanto ne sapevamo prima.
Se l’energia necessaria per ogni contrazione muscolare viene in gran parte dalla glicogenolisi/glicolisi (si parla sempre di sforzi di durata a bassa intensità), non si è certi in che percentuale questa soddisfa l’intero processo; è il 90%, l’80% o il 65%? Non è possibile saperlo. Il resto dell’energia proviene dal sistema della fosfocreatina e dal metabolismo aerobico.
Ma la necessità energetica della cellula muscolare non si esaurisce nella contrazione; infatti, durante la fase di rilassamento occorre ATP per risintetizzare il glicogeno (l’abbiamo visto sopra), per mantenere l’omeostasi, per i processi di ripolarizzazione, per il funzionamento delle pompe, per ripristinare una parte della fosfocreatina, ecc. È probabilmente in questa fase che il metabolismo aerobico ha un peso significativo (rispetto a quello anaerobico) e venga ossidato gran parte del lattato formato durante la contrazione (Brooks 2020).
A mio parere, i risvolti applicativi più importanti riguardano il fatto che esistono 2 fasi ben distinte, cioè la contrazione e il rilassamento muscolare.
Ad esempio, prima si era portati a pensare che il sistema della fosfocreatina entrasse in funzione solamente oltre una certa potenza o nei primi secondi dello sforzo; adesso sappiamo come questo metabolismo (anaerobico alattacido) entra in funzione anche nelle intensità più basse, nei primi millisecondi della contrazione muscolare; in altre parole, il turnover della fosfocreatina in passato è stato sempre sottostimato. Che ricadute applicative possono avere queste nuove conoscenze? Lo vedremo nei prossimi paragrafi.
Ma cosa succede in sforzi brevi ed intensi?
La teoria dello shuttle del lattato è fondata su studi effettuati tramite sforzi di bassa intensità e lunga durata, proprio per analizzare quale fosse il comportamento dei metabolismi in uno stato stazionario. Non sono ancora disponibili indagini con RMN con campionamento al ms su impegni muscolari ad alta intensità, o comunque protratti fino ad esaurimento.
Confrontando il vecchio modello dei metabolismi energetici (che non è errato, ma fornisce una versione macroscopica di quello che accade) con quello dello shunt del glicogeno (che fornisce una visione molto più dettagliata), è ragionevole intuire come man mano che prosegue lo sforzo di alta intensità, la concentrazione di fosfocreatina scenda progressivamente con un andamento comunque oscillatorio (a seconda che la fibra muscolare si trovi in stato di contrazione o rilassamento), ed aumenti sempre di più il contributo della glicogenolisi/glicolisi ed in parte del metabolismo aerobico. Ad un certo punto comunque, la fatica (che ricordiamo ha cause multifattoriali) non permetterà di mantenere un’intensità massimale, facendo così calare la potenza prodotta dal muscolo. Questo è quello che potrebbe avvenire in una gara di 100m.

Quello che è importante comprendere, è che l’insorgenza della fatica non avviene per l’inabilità dei metabolismi di produrre energia (cioè ATP); non a caso, anche in sforzi atletici intensi, l’ATP rimane costante (Westerbland et al 2002), ed anche quando cala (come in contrazioni di muscolo quasi isolato, come può essere una leg extension), la fatica compare quando la sua concentrazione è comunque costante (Layec et al 2015).
La fatica è un meccanismo protettivo della cellula e dell’organismo che evita che la loro integrità venga compromessa; è un insieme di segnali che inibisce l’entità ed il rendimento della contrazione muscolare.
Dai dati disponibili in bibliografia internazionale, quello della fosfocreatina è l’unico dei sistemi energetici che in alcune condizioni può andare in difficoltà (Walter et al 1999); infatti, in sforzi particolarmente intensi la sua concentrazione può quasi azzerarsi, ma in questi casi gli altri 2 metabolismi riescono a mantenere la concentrazione cellulare di ATP costante. Questo testimonia come lavorino in maniera integrata, per garantire la produzione di energia dai primissimi istanti del movimento, fino a sforzi di lunga durata (Baker et al 2010).
Andiamo ora a fare un breve approfondimento delle 2 molecole chiave dei metabolismi energetici (e della teoria dello shunt del glicogeno), cioè il lattato ed il glicogeno; ci permetterà di estrapolare le ricadute applicative in chiave metodologia d’allenamento e nutrizione.
“Il lattato come fulcro del metabolismo”
È il titolo dell’ultima revisione di Brooks del 2020, che segue quella altrettanto importante del 2018; una volta si credeva che la cellula ricorresse alla produzione del lattato quando non era presente all’interno una quantità di ossigeno sufficiente per ossidare il piruvato (prodotto finale della glicolisi); oggi sappiamo che il lattato viene prodotto anche in presenza di elevate quantità di ossigeno all’interno della fibra, con un rapporto tra lattato/piruvato superiore a 500, quando si è sottosforzo. Questo dimostra come durante l’istante della contrazione muscolare sia necessaria un’elevata produzione di ATP grazie alla glicogenolisi/glicolisi. Il lattato prodotto può seguire diverse vie:
- Essere ossidato a scopo energetico nel mitocondrio della stessa fibra che l’ha prodotto: lo scopo è quello di sostenere una parte minore della contrazione muscolare, fornire l’energia per la risintesi di glicogeno (dal glucosio) durante la fase di rilassamento, mantenere l’omeostasi cellulare, mantenere il funzionamento delle pompe, per la ripolarizzazione, ecc.
- Migrare nelle fibre muscolari attigue e svolgere le stesse funzioni: si ipotizza che il destino di più della metà del lattato prodotto segua la destinazione di questi primi 2 punti. Sembra in particolar modo che il lattato (quando passa da fibra a fibra), vada dalle veloci verso le lente (Park et al 2015).
- Venga ossidato in altri organi: in particolar modo sottosforzo, il cuore utilizza il lattato come fonte primaria di energia (preferendolo al glucosio). Non a caso è una strategia estremamente vantaggiosa, in quanto il lattato viene prodotto in grandi quantità dal muscolo, mentre il glucosio è una fonte importante per risintetizzare il glicogeno muscolare. Inoltre, il lattato è in grado di diffondersi più velocemente nel corpo rispetto al glucosio. In misura inferiore, anche il cervello, l’intestino, polmoni ed altri organi utilizzano il lattato a scopo energetico.
- Viene utilizzato per la neoglucogenesi, cioè viene trasformato in glucosio; una volta si credeva che questo avvenisse principalmente nel fegato, grazie al Ciclo di Cori. Oggi sappiamo che anche il rene svolge questa funzione; il 50% del glucosio formato dal lattato, proviene dai reni (Cano 2002). La neoglucogenesi è estremamente importante (la approfondiremo meglio successivamente), perché permette di risintetizzare le unità fondamentali del glicogeno, cioè il glucosio.
- Sintesi diretta di glicogeno nelle fibre: malgrado ancora non si conosca con precisione la via metabolica, in diversi studi è stato visto come il lattato, all’interno delle cellule muscolari (soprattutto quelle veloci) può contribuire a sintetizzare unità glicosidiche del glicogeno (McLane et al 1979, Johnson et al 1985, Bonen et al 1990, McDermott 1992, Bangbo et al 1997). In media dal 13 al 27% del lattato prodotto dalle fibre muscolari viene riconvertito direttamente in glicogeno (Bangsbo et al 1991). Questo rappresenta un vantaggio non indifferente, in quanto permette di risintetizzare velocemente glicogeno senza dover utilizzare il glucosio.
- Ultima funzione, che però non è energetica, è regolatoria: il lattato rappresenta un importante segnale in grado attivare reazioni all’interno della cellula, nelle cellule attigue e in organi e tessuti lontani, compresa la sintesi di ormoni. Non a caso, è anche definito “lactormone”.
Il lattato non determina l’acidosi, e nemmeno la fatica
Per moti anni si è creduto che il prodotto finale della glicolisi anaerobica fosse l’acido lattico; si ipotizzava che questo si dissociasse in lattato + H+ (idrogenione) il quale contribuisse ad abbassare il ph, incidendo sulla fatica. Oggi sappiamo che non è così.

Sopra potete vedere la reazione ultima della glicolisi anaerobica; il primo punto importante, è che il risultato finale non è l’acido lattico, ma il lattato (cioè l’acido lattico senza l’H+). Non solo, nella stessa reazione è possibile vedere come nella parte destra della formula (cioè quando il piruvato si trasforma in lattato) scompaia lo ione H+ presente nella parte sinistra, contribuendo a ridurre l’acidosi.
Ora, non si sa con certezza se in altre reazioni della glicolisi si producano H+ che vadano ad aumentare l’acidosi, ma non è possibile attribuire al lattato questa “colpa” (Brooks 2018).
Quindi, malgrado si possa essere certi che un abbassamento del ph (cioè un aumento dell’acidosi) oltre un certo livello possa far insorgere la fatica, attualmente non si sa quali siano le reazioni metaboliche che influiscano maggiormente su questo fenomeno. Non solo, malgrado il piruvato sia “accompagnato” da un protone (H+), si è certi che questo metabolita più di tanto non si accumula nella cellula, visto che il rapporto lattato/piruvato passa da un valore di 10 a riposo, ad oltre 500 sottosforzo.
Quindi, la prossima volta che qualcuno vi dice “ho male alle gambe perché ho ancora l’acido lattico da ieri…”, fategli capire che il lattato non è “il colpevole” del mal di gambe; non solo, anche quando la sua concentrazione diventa molto alta, già dopo 30’ la fine dello sforzo questa si dimezza.
La sensazione del mal di gambe tardivo (cioè quello del giorno dopo) è dovuto alla stimolazione dei recettori del dolore, ad opera di vari metaboliti delle cellule del sistema immunitario migrate verso l’area muscolare, che durante lo sforzo ha subito microlesioni alle fibre, e/o alla fuoriuscita di materiale fibrillare.
Acido lattico, intestino e performance: il caso Veillonella
La Veillonella è un batterio presente nell’apparato digerente, ma si può trovare anche il quello respiratorio; fa parte del microbiota, cioè di quell’insieme di batteri, virus e funghi che vivono in simbiosi con il nostro organismo. Malgrado si stia sempre più comprendendo l’importanza dell’interazione tra il nostro organismo ed il nostro microbiota, poco si conosce dell’interazione specifica delle varie specie.
Il “caso Veillonella” è un tipico esempio di come possa esserci correlazione tra composizione del microbioma e performance, ma allo stesso tempo sono necessari ancora diversi studi per approfondire eventuali rapporti di causa/effetto. Essendo presenti un numero limitato di ricerche (ma molto interessanti), vediamo di riassumerle brevemente e trarne i possibili riscontri pratici: in uno studio del 2019 (Sheiman et al 2019) tramite l’osservazione del microbioma (cioè l’analisi del DNA dei batteri dell’intestino) venne visto come questo batterio fosse in misura maggiore dopo una maratona, rispetto a 7 giorni prima; gli stessi risultati furono poi confermati da uno studio seguente.
Precedentemente, fu visto come la Veillonella è in grado di trasformare il lattato in 2 acidi grassi a catena corta (acetato e propionato) grazie alla via metabolica del Metilmalonil-coenzima A (Stephen et al 1973), la cui attività è stata vista incrementare proprio sottosforzo. L’acetato è in grado di entrare nel ciclo di Krebs fornendo energia per via ossidativa, ma è il propionato ad aver attirato l’interesse dei ricercatori.
Infatti, questa molecola può entrare nella neoglucogenesi, cioè portare alla neoformazione di glucosio, che abbiamo visto precedentemente essere molto importante per la riformazione del glicogeno durante lo sforzo. Ma lo studio di Sheiman et al 2019 non finisce qui; infatti gli autori cercarono di verificare se un aumento della presenza di Veillonella (tramite introduzione) fosse in grado di promuovere questo fenomeno, migliorando la resistenza di topi di laboratorio. I risultati furono che i topi a cui fu inoculato questo batterio migliorarono del 13% la loro resistenza in un test ad esaurimento, con una minore infiammazione post-sforzo rispetto alle altre cavie.

Bene, a questo punto, tutti ad utilizzare integratori di probiotici con Veillonella (che ovviamente non esistono) per andare più forte!
Naturalmente sto scherzando, anche perché mancano ancora conferme di studi su umani; non solo, affinchè i risultati di uno studio possano essere considerati validi, è necessaria la conferma di altre ricerche.
Infatti, malgrado la Veillonella sembra (non tutti gli studi lo confermano) maggiormente presente negli sportivi rispetto ai sedentari (Petersen et al 2017), non è l’unico batterio ad avere questo effetto sui topi (Okamodo et al 2019); anche il Kefir (che rappresenta un insieme di diversi batteri) ha dimostrato effetti simili (Hsu et al 2018).
Quello che è importante capire, è che il nostro microbioma si adatta all’allenamento, modificando l’attività e le specie presenti al fine di facilitare l’adattamento allo sforzo di endurance. Con tutta probabilità, vengono maggiormente espresse quelle specie che permettono di metabolizzare il lattato in altri substrati più velocemente utilizzabili a scopo energetico (come l’acetato) o con caratteristiche glucogenetiche (come il proprionato). Ricordo che anche l’intestino ha proprietà glucogenetiche (Mithieux et al 2014), cioè è in grado di convertire specifici metaboliti (come il proprionato) in glucosio.
Quello che ancora non si conosce, e se l’integrazione alimentare con tali specie possa favorire o meno la crescita di questi batteri nell’intestino e di conseguenza la performance; non solo, in uno studio (Morita et al 2023) è stato visto come le alfa-ciclodestrine (che sono delle fibre, dei pre-biotici) sono in grado di incrementare la popolazione di una specie batterica (Bacteroides uniformis) potenzialmente in grado di migliorare la performance di endurance grazie alla produzione di acidi grassi a corta catena. Come più volte ripetuto, uno studio non può fare la scienza, ma servono tante ricerche (possibilmente da laboratori differenti) affinchè si possa affermare che una qualsiasi sostanza possa migliorare la performance.
Quello che attualmente invece è stato compreso, è che la salute del microbioma può incidere sulla performance sportiva e sul recupero. Per ulteriori dettagli, potete leggere il nostro post su Sport e Microbioma.
Glucosio, integratori e stimoli allenanti
Nel primo capitolo abbiamo visto come il glicogeno fosse la fonte energetica primaria per la fase di contrazione muscolare e come la cellula faccia di tutto per ripristinarlo durante la fase di rilassamento. È quindi presumibile ipotizzare, come la maggior parte del glucosio che entra nella fibra muscolare non venga direttamente metabolizzato dalla glicolisi, ma vada a formare le unità glicosidiche del glicogeno.
Considerando che a riposo nel sangue siano presenti solo 5 grammi di glucosio, è evidente come la fisiologia dell’organismo sia funzionale nell’accumulare questa molecola in altri organi (fegato), a risparmiarlo a scopo energetico durante lo sforzo (facendo preferire il lattato da altri organi particolarmente attivi come cuore e cervello), a favorirne la sintesi da altre molecole (neuglucogenesi) e a mettere in condizioni l’apparato digerente di assorbirlo velocemente al fine di veicolarlo verso i muscoli.
Ma partiamo proprio da quest’ultimo punto, sul quale studi e ricerche da anni cercano di dare una risposta alle seguenti domande:
- Quanti carboidrati è in grado di assorbire sottosforzo l’intestino?
- Qual è la formulazione migliore che permette una più veloce digestione ed assorbimento?
Attualmente, è possibile affermare che la quota ideale per massimizzare la performance in sforzi particolarmente prolungati sia quella di ingerire una quota di circa 80 grammi di carboidrati ogni ora, con un range compreso tra 70-90g (Guillochon et al 2017, Smith et al 2013, King et al 2018).
La formulazione ideale prevede i carboidrati suddivisi circa la metà (o qualcosa meno) di fruttosio e l’altra metà (o qualcosa di più) in maltodestrine (O’Brien et al 2013). Questo, perché utilizzando carboidrati diversi si sfruttano differenti meccanismi di assorbimento, di conseguenza si riesce ad assimilarne una quantità maggiore.

Questi dati sono estrapolati da studi effettuati in laboratorio con cicloergometro, perché permette di protrarre più a lungo lo sforzo (rispetto alla corsa, ad esempio). Non solo, non è raro che in sport in cui è presente uno “sballottamento” dell’apparato digerente (come la corsa), sia più facile andare incontro a problematiche di natura intestinale, anche ai dosaggi indicati sopra.
Tutto questo per far capire come dalla teoria alla pratica possa esserci una certa differenza. Quello che è importante, è capire quali possano essere le condizioni ideali per ogni atleta, considerando sia le indicazioni provenienti dalla bibliografia internazionale che la soggettività dell’individuo, che lo sport praticato.

Nel nostro articolo dedicato all’idratazione ed integrazione, potete trovare tutte le indicazioni su questo argomento, considerando anche l’acqua (per l’idratazione) ed eventualmente i Sali. Per comprendere invece come limitare disturbi gastrointestinali, vi invitiamo a leggere il capitolo Train the gut (allena l’intestino) a questo articolo. Ma facciamo qualche passo avanti, e vediamo quali possano essere (come abbiamo fatto per la Veillonella) le ricadute applicative in base a quelle che sono le scoperte più recenti.
L’Hydrogel e le ciclodestrine sono il futuro dell’integrazione in gara?
Se parliamo di marginal gains associati al metabolismo dei carboidrati, allora non possiamo non citare l’hydrogel e le ciclodestrine altamente ramificate.
L’hydrogel è una normale formulazione di carboidrati a cui viene aggiunto alginato di pectina e sodio; il risultato è che abbandona più velocemente lo stomaco perché forma un gel (al ph dello stomaco) che “nasconde” i carboidrati ai recettori del duodeno. Questi recettori infatti, si trovano all’inizio dell’intestino, appena fuori dallo stomaco; sono responsabili del rallentamento della velocità di svuotamento gastrico (cioè dello stomaco) quando vengono a contatto con dei carboidrati (glucosio, maltodestrine, ecc.).
Il gel che si viene a creare grazie alla presenza di alginato di pectina, “nasconde” i carboidrati presenti nel prodotto facilitando lo svuotamento gastrico; il risultato principale, è quello di limitare notevolmente il rischio di problematiche intestinali. Non solo, permetterebbe ipoteticamente di poter ingerire una quota superiore di questi macronutrienti che, in sforzi superiori alle 2h30’, può influenzare statisticamente la performance.
Le ciclodestrine altamente ramificate (HBCD) hanno benefici sovrapponibili (cioè quello di ridurre l’insorgenza di problematiche intestinali), ma sfruttano un meccanismo leggermente diverso. Hanno sempre una maggiore velocità di svuotamento gastrico rispetto ai carboidrati più comunemente utilizzati (cioè le maltodestrine), ma lo ottengono grazie ad un peso molecolare molto superiore, che riduce l’osmolarità e di conseguenza facilita lo svuotamento dello stomaco.
Potete approfondire ulteriormente (oltre a trovare i migliori prodotti) leggendo il nostro articolo su hydrogel e ciclodestrine.
Chiedere sempre consiglio a personale qualificato prima di assumere qualsiasi tipo di integratori.
Come allenare l’organismo ad utilizzare substrati alternativi al glucosio?
Abbiamo visto che il glucosio è una molecola fondamentale per ripristinare le scorte di glicogeno durante il rilassamento della fibra muscolare. Se è importante comprendere quale sia la migliore strategia per integrare al meglio in gara, è fondamentale conoscere anche altri 3 aspetti importanti relativi al metabolismo dei carboidrati, soprattutto per gli sport di endurance:
- Come stoccare nei muscoli e nel fegato la massima quantità di glicogeno possibile prima della gara (senza andare incontro ad effetti avversi).
- Come allenare (nel caso in cui sia possibile) nel miglior modo possibile la gluconeogenesi
- Come abituare l’organismo ad utilizzare substrati alternativi (in particolar modo lipidi) ai carboidrati per risparmiare glucosio e glicogeno,
Sul primo punto non ci dilunghiamo, in quanto è stato già ben approfondito nel nostro post dedicato al carico di carboidrati. Andiamo invece a vedere quali sono i risultati degli studi più recenti sul secondo e sul terzo punto.
È allenabile la neoglucogenesi?
Ora sappiamo quanto sia importante la sintesi di glucosio a partire da altri substrati; ma dove avviene? Quali sono i substrati principali?
Già nei capitoli precedenti, abbiamo visto come questa via metabolica fosse presente prevalentemente nel fegato e nei reni, ed in misura minore nell’intestino e nel cervello (Cano 2002), ma vediamo in maniera più esaustiva quali sono i substrati principali della neoglucognesi durante l’attività fisica:
- Lattato: sottosforzo è sicuramente il principale, sia in virtù dell’elevata produzione durante la contrazione muscolare, che dovuto al fatto che si diffonde facilmente negli organi e tessuti.
- Aminoacidi glucogenetici: questi possono arrivare al fegato e seguire la via metabolica della formazione di glucosio. L’aminoacido più utilizzato è l’alanina, tramite il ciclo alanina-glucosio. Questa via metabolica è molto importante, perché prevede l’interazione degli aminoacidi con il piruvato prodotto dalla glicogenolisi/glicolisi. Altro aminoacido molto utilizzato nella gluconeogenesi è la glutammina, che segue un meccanismo molto simile a quello dell’alanina. Altri aminoacidi contribuiscono in misura minore alla formazione di glucosio.
- Glicerolo: l’utilizzo dei trigliceridi a scopo energetico, prevede la scomposizione di questi in acidi grassi più glicerolo. Se gli acidi grassi vengono legati al CoA per poi seguire la via dell’ossidazione, il glicerolo viene utilizzato per formare glucosio.
- Propionato: è un acido grasso a corta catena, prodotto prevalentemente all’interno dell’intestino (vedi Veillonella). Insieme all’Acetato rappresenta il substrato più significativo per la neoglucogensi che proviene dall’intestino; non solo, a mio parere rappresenta un ambito di margine di guadagno prestativo ancora poco conosciuto. Per approfondire, potete leggere l’articolo sulle alfa-ciclodestrine.

Ricordo che anche il glicogeno presente nel fegato (75-100g in condizioni di riposo) contribuisce a rilasciare glucosio che poi finirà nel sangue; per questo motivo, il fegato ha una doppia importanza, cioè quella di rilasciare glucosio stoccato al suo interno (sottoforma di glicogeno) e di neoformare glucosio da altri metaboliti (lattato, glicerolo ed aminoacidi).
Ma torniamo al titolo di questo paragrafo: quello che a mio parere è importante comprendere, è se alcune forme d’allenamento (rispetto ad altre) sono in grado di stimolare questa importante via metabolica o se modificazioni di natura alimentare possono stimolarla più di altre.
Quello che attualmente sappiamo dalla bibliografia internazionale, è che la neoglucogenesi sottosforzo è 3 volte più efficace nei soggetti allenati rispetto ai sedentari (Bergman et al 2000); in ogni modo, la manipolazione dietetica (cioè modificare proporzione tra grassi e carboidrati) non influenza la neoglucogenesi (Webster et al 2016). Piuttosto, gli stessi autori trovarono come aumentando la quota calorica dei grassi, e riducendo quella dei carboidrati (ciclisti allenati), veniva parzialmente incrementato l’utilizzo dei lipidi a scopo energetico. Questo concetto è di elevatissimo interesse all’atto pratico, perché apre le porte a sviluppi metodologici abbinati a modificazioni dietetiche. Vediamo meglio questo concetto nel prossimo capitolo.
Come consumare più grassi a scopo energetico modificando l’apporto alimentare
Nel precedente capitolo abbiamo visto come aumentando l’apporto calorico dei lipidi e diminuendo quello di carboidrati (mantenendo comunque costanti le calorie totali) si assiste ad un leggero “spostamento” del metabolismo energetico verso i grassi (Psilander et al 2013, Bartlett et al 2015 e Webster et al 2016). Questo concetto potrebbe rivelarsi interessante in quelle discipline di lunga durata (maratone, trail, corse ciclistiche, triathlon di una certa durata, ecc.) in cui è importante “risparmiare” le scorte glucidiche nella prima parte di gara. Malgrado questo, all’atto pratico è stato visto come diete iperlipidiche (compresa la dieta ketogenetica) fossero deleterie per chi pratica sport di endurance, perché limitano l’apporto di carboidrati e di conseguenza impoveriscono le scorte di glicogeno, necessarie per mantenere i ritmi elevati anche nelle gare lunghe; non solo riducono anche la capacità dell’organismo di assorbire velocemente e metabolizzare i carboidrati (Burke et al 2017, Helge et al 2017 e Burke 2020).
La stessa cosa si potrebbe dire per l’esercizio fisico in caso di digiuno pre-sforzo (mantenendo normale l’apporto calorico giornaliero); anche la pratica sportiva in queste condizioni è stata visto aumentare il consumo dei lipidi intracellulari, con un effetto di risparmio del glicogeno a basse intensità (Murray et al 2018, Vicente-Salar 2015), ma è evidente come questa condizione non possa essere l’ideale in situazione di gara, in cui è fondamentale la necessità di avere elevate scorte di glicogeno.
Quello che emerge in questi studi, è comunque interessante; infatti, le ricerche sopracitate evidenziano i risultati deleteri di settimane dello stesso approccio dietetico.
Ma può essere proficuo adottare queste strategie saltuariamente e solo in alcune condizioni?
Autori particolarmente esperti nel settore come Burke 2015 affermano come non sia da escludere, in un ambito di individualizzazione dei protocolli, modificare in alcune giornate la dieta e l’allenamento al fine di abituare l’organismo a massimizzare l’utilizzo dei grassi a basse intensità.
È il classico esempio del Training Low – Compete High, che abbiamo visto anche nell’approccio dietetico alla maratona. Fondamentalmente si tratta di ridurre l’apporto di carboidrati per un numero limitato di pasti (1, al massimo 2) ed eseguire l’allenamento successivo con lo scopo di abituare l’organismo ad esercitarsi con basse scorte di glicogeno, utilizzando ritmi comunque non elevati per evitare di arrivare all’esaurimento.

Bartlett et al 2015 confermarono come in queste condizioni vengano enfatizzati i segnali cellulari responsabili dell’adattamento dell’ossidazione dei substrati energetici; gli autori concludono che per evitare che questi protocolli comportino effetti deleteri sul metabolismo dei glucidi, andrebbero inseriti saltuariamente, mantenendo un adeguato apporto di carboidrati nella maggior parte delle giornate ed un apporto elevato nei giorni che precedono le gare o durante le gare multistage (esempio corse a tappe). All’interno di questo gruppo di ricerca, troviamo anche James Morton, che in passato si è occupato dell’aspetto nutrizionale e metabolico del Team Sky di Ciclismo e del Liverpool FC.
In questo suo interessante articolo sintetizza alcune strategie nutrizionali attuate al Team Sky (ora Team Ineos); quello che sottolinea (confermando quanto appena scritto), è che allenandosi a basse intensità e con una riduzione dell’apporto dietetico di carboidrati, si massimizza l’utilizzo di substrati alternativi ai glucidi. Quello che è difficile, è il comprendere la giusta dose di carboidrati necessari (per non abbassare troppo i livelli di glicogeno) e l’intensità ideale da mantenere affinchè non ci sia un esaurimento eccessivo delle scorte glucidiche; tutto questo va inoltre personalizzato per ogni atleta, che deve essere seguito giornalmente anche da personale medico. Non a caso, questi protocolli venivano fatti esclusivamente nelle gare a tappe o nei ritiri (quando era presente tutto il personale).
Ma che spunti può trarne la maggior parte degli atleti o dei professionisti del settore?
A mio parere i punti, in ordine di importanza, sono i seguenti:
- Segui uno stile di vita adeguato, curando anche il recupero nel caso ti alleni 5 o più volte a settimana.
- Segui una dieta varia ed equilibrata, assicurandoti giornalmente una quota calorica, glucidica e proteica adeguata alle tue necessità; cura particolarmente l’apporto di frutta e verdura di stagione. Se hai dei dubbi, rivolgiti ad un nutrizionista o un dietologo.
Nel caso in cui tu voglia provare la strategia “Training low”, per discipline di endurance della durata superiore a 1h30’-2h:
- Utilizzala non più di una volta alla settimana, preferendo ritmi bassi e una durata dell’allenamento non elevata. Questo ti permetterà di comprendere al meglio come individualizzare in futuro questo tipo di strategia.
- Modifica l’apporto dietetico nel pasto che precede l’allenamento “Training low” in maniera estremamente graduale.
- Ricordati che arrivare a fine allenamento affaticato a tal punto da non tenere le basse intensità, significa che hai esagerato con la riduzione dei carboidrati o la durata della seduta.
- “Raschiare il barile” troppe volte significa andare incontro ad affaticamenti che tendono a limitare i miglioramenti della condizione (sovraffaticamento o sovrallenamento).
- Prima degli allenamenti più impegnativi (per intensità e/o durata) segui una dieta con un normale (o superiore) apporto di carboidrati, altrimenti non riuscirai a tenere le andature per i tempi indicati. Se necessario (ad esempio nei lunghi per i maratoneti), integra durante la seduta per abituare l’intestino a quello che dovrà poi ingerire in gara.
- Puoi considerare il “digiuno pre-allenamento” analogamente alla seduta “Training-low”, ma con le stesse raccomandazioni di sopra (gradualità, intensità e durata moderata della seduta, ecc.). Questa strategia prevede comunque lo stesso apporto calorico giornaliero e di nutrienti di una dieta normale; quello che cambia sono i momenti della giornata in cui ci si alimenta. La strategia ideale, più semplice e più comune è l’allenamento mattiniero a digiuno.
Come potete ben vedere, le raccomandazioni nell’utilizzo della strategia “Train-low” sono diverse, proprio perché può essere un’arma a doppio taglio, visto la necessità di individualizzarla; per chi è interessato, è consigliabile effettuarla facendosi seguire da personale qualificato e solo se si praticano discipline di endurance con competizioni della durata superiore ai 75-90’.
Quello che è importante tenere a mente, è che di norma un allenamento adeguato, una dieta equilibrata ed un corretto stile di vita sono i punti cardine per allenarsi e gareggiare al meglio.
Fino ad ora abbiamo visto le ricadute applicative della teoria dello shunt del glicogeno a livello metabolico ed alimentare; ora facciamo un breve excursus di quelli che possono essere le ricadute applicative a livello di metodologia d’allenamento.
Teoria dello Shunt del glicogeno: Verkhoshansky l’aveva capita molto prima
Il titolo non vuole essere provocatorio, ma ricordare Yuri Verkhoshansky che dal 1995 divenne consulente del CONI, contribuendo, con le sue esperienze e le sue competenze, alla crescita della conoscenza del processo di allenamento in tantissimi sport, soprattutto nell’atletica leggera.
Da laureato in Scienze Motorie ed appassionato di atletica leggera (fondo e mezzofondo), mi sono sempre interessato ai suoi metodi finalizzati allo sviluppo della potenza aerobica per mezzofondisti; metodi non sempre facili da comprendere e da “digerire”, perché particolarmente innovativi…ma che hanno anticipato i tempi, e sono tutt’oggi più che attuali, proprio perché trovano il razionale nella teoria dello Shunt del Glicogeno, che allora ovviamente non era conosciuta. Ma andiamo per ordine.
Nei primi capitoli di questo articolo abbiamo visto come nel definire l’attivazione dei vari metabolismi non è solo l’intensità dello sforzo, ma anche il rapporto tra il tempo di contrazione e rilassamento nel singolo gesto motorio. Nella pubblicazione 1 del 2003 di AtleticaStudi, Verkhoshansky presentò la sua metodologia dei balzi nel sistema di preparazione nell’atletica leggera. Una delle evidenze che mi colpì di questo metodo fu il “miglioramento della capacità dei muscoli di rilassarsi istantaneamente dopo l’esecuzione di sforzi motori potenti”.
Praticamente l’autore proponeva il miglioramento della massima potenza aerobica anche tramite l’utilizzo di corsa balzata in salita con stacchi avanti-alto cercando di fare balzi lunghi e potenti. Ogni ripetizione era formata da soli 6-8 balzi, organizzati in serie con recuperi anche superiori ai 2-3’ tra ogni serie. Ovviamente non mi dilungo sui vari mezzi allenanti proposti, ma riporto sotto il motivo per cui l’esecuzione richiesta era quella di balzi potenti, anziché una normale corsa veloce massimale. Leggete attentamente queste sue 3 frasi sotto, riportate sulla rivista Teknosport (oggi non più prodotta).
Un’elevata resistenza esterna con una frequenza moderata dei movimenti sviluppa prevalentemente il meccanismo del creatin-fosfato e la potenza aerobica con una scarsa sollecitazione della glicolisi.
Il fattore principale non è la velocità della corsa, ma l’accentuata energica, azione della gamba di spinta accompagnata da un’azione accentuata di avanzamento della coscia dell’arto libero. Così si evitano sia il debito di ossigeno che l’aumento della concentrazione del lattato ematico e muscolare, e quindi gli atleti possono usare un volume relativamente elevato di lavoro. Questo tipo di lavoro aumenta le capacità anaerobiche […] e le capacità ossidative dei muscoli
Un lavoro con ritmo elevato e con una scarsa resistenza esterna provocherebbe invece un dispendio energetico eccessivo ed, in buona sostanza, diventerebbe inutile.
Sostanzialmente non solo “anticipa” l’importanza degli sforzi brevi ed intensi a supporto dell’allenamento per sport di endurance, oggi meglio conosciuto nel contesto dell’allenamento polarizzato; ma viene posto l’accento sul rapporto tra il tempo di contrazione dell’arto in appoggio (che deve essere minimo) e il tempo di rilassamento (che deve essere massimo), al fine di ridurre la spesa energetica a carico della glicolisi, risparmiando quindi glicogeno; tutto questo riguarda ovviamente i mezzi allenanti costituiti da balzi, da utilizzare solamente da atleti con adeguate caratteristiche neuromuscolari. Questi concetti venivano esposti senza essere a conoscenza degli studi che diedero origine teoria dello Shunt del Glicogeno.
Teoria dello shunt del glicogeno e sport di squadra
Nelle discipline in cui la componente tecnico-tattica riveste un ruolo fondamentale, i movimenti ed i gesti atletici sono estremamente più vari rispetto agli sport ciclici. Nel nostro post dedicato alla Potenza Aerobica nel calcio, abbiamo confrontato l’espressione della potenza aerobica tra dilettanti e professionisti; il risultato è stato che i professionisti non necessariamente presentano una maggior velocità aerobica lineare, ma sono dotati di una migliore efficienza energetica nella situazione di gioco e nei cambi di direzione.
Questo si spiega semplicemente dal fatto che un maggior livello di coordinazione nei gesti specifici (rispetto ai dilettanti) permette di ridurre i tempi di contrazione e reclutare selettivamente i muscoli che permettono l’esecuzione del gesto; il risultato è l’utilizzo di una quantità inferiore di glicogeno (tempi di contrazione ridotti) ad ogni contrazione ed un utilizzo di meno fibre muscolari grazie ad una maggior maestria motoria. Tutto questo si traduce in una miglior potenza aerobica applicata al gioco ed una maggiore resistenza alla fatica; per approfondire gli aspetti metodologici potete leggere il nostro post Quale potenza aerobica nel calcio.
Conclusioni
Siamo arrivati al termine di questo lungo articolo, in cui ho cercato di approfondire come la teoria dello shunt del glicogeno abbia migliorato le conoscenze dei metabolismi energetici, e le applicazioni pratiche che ne derivano.
Credo sia veramente importante rimanere continuamente aggiornati sulla propria disciplina di competenza, ma allo stesso tempo ampliare anche le conoscenze di quelle affini, come può essere la comprensione dettagliata dei metabolismi per chi opera nella preparazione dei vari sport. Questo permette di sviluppare più abilità conoscitive e di sfruttare il pensiero laterale; per approfondire cosa sia il pensiero laterale, vi consiglio i libri di Edward De Bono.
Oggi l’accesso al sapere ed alla conoscenza è estremamente facilitato rispetto a 20-30 anni fa, quindi per fare la differenza nel proprio campo è necessario più che mai utilizzare la creatività e l’intuito, padroneggiando non solo le competenze specifiche del proprio sport, ma anche quelle affini.
Per chi opera nel calcio, cliccando sull’immagine sotto potete accedere al post (continuamente aggiornato) in cui elenco i testi che attualmente ritengo più utili per chi lavora in questo settore.

Bene spero che l’articolo ti sia piaciuto e ti sia stato utile; se vuoi rimanere aggiornato sulle nostre pubblicazioni e revisioni, connettiti al mio profilo linkedin.
Autore dell’articolo: Melli Luca, preparatore atletico AC Sorbolo, istruttore Scuola Calcio A.S.D. Monticelli Terme 1960 ed Istruttore di Atletica leggera GS Toccalmatto. Email: melsh76@libero.it








Comments are closed